2 de julio de 2010
ESTRUCTURA FLORAL
Las estructuras reproductoras de las angiospermas se agrupan en órganos llamados flores, típicas del grupo. Cada flor es, por lo tanto, un órgano complejo formado por diversas partes, que alberga, protegiéndolos, al gametofito masculino y al femenino.
Es un tallo de crecimiento definido, con entrenudos muy cortos, en el que se insertan hojas modificadas, los antófilos u hojas florales.
Se puede considerar a la flor como un eje complejo reducido en el curso de la evolución y derivado de ancestros gimnospérmicos, ya que la vascularización del eje floral es más compleja que el de una rama simple, y porta órganos laterales que no son homólogos de hojas (ovario, óvulos).
Es un tallo de crecimiento definido, con entrenudos muy cortos, en el que se insertan hojas modificadas, los antófilos u hojas florales.
Se puede considerar a la flor como un eje complejo reducido en el curso de la evolución y derivado de ancestros gimnospérmicos, ya que la vascularización del eje floral es más compleja que el de una rama simple, y porta órganos laterales que no son homólogos de hojas (ovario, óvulos).
Tamando una flor se puede encontrar en ella una serie de verticilos superpuestos de órganos laterales. Desde la base del pedicelo floral hacia arriba los verticilos son:
El Cáliz, formado por elementos llamados sépalos.
La Corola, formada por elementos llamados pétalos.
El Androceo, formado por elementos llamados estambres.
El Gineceo, formado por elementos llamados carperlos.

Una flor es completa cuando está formada por todos los verticilos, y es incompleta cuando carece de alguno de ellos.
Las flores de las angiospermas poseen una envoltura floral, un perianto, el cual esta constituído por los verticilos de la flor: el cáliz y la corola. Si ambos ciclos se diferencian entre sí la flor es heteroclamídea. A veces la forma y el color de estos dos verticilos son semejantes, en este caso se habla de perigonio y la flor es homoclamídea. Sus piezas florales reciben el nombre de tépalos. Si la flor carece de perianto recibe el nombre de flor desnuda, aclamídea o aperiantada. Sin embargo, si sólo carece de cáliz recibe el nombre asépala, y si carece de corola es apétala.
El Androceo constituye el verticilo que esta asociado con el origen del gametofito masculino. Está intercalado entre la corola y el ovario cuando la flor es completa. Cada unidad del androceo es un estambre.
Los estambres pueden estar libres, recibiendo el nombre dialistémonos o diversamente unidos. Según como se encuentran unidos, tendremos:
Androceo monadelfo: los estambres están soldados entre sí por sus filamentos constituyen un sólo grupo.
Androceo diadelfo: los estambres están soldados entre sí por sus filamentos constituyendo dos grupos.
Androceo poliadelfo: los estambres están soldados entre sí por sus filametos formando varios grupos.
Los estambres que no producen polen fértil reciben el nobre de estaminodios.
25 de junio de 2010

Estructuras reproductoras
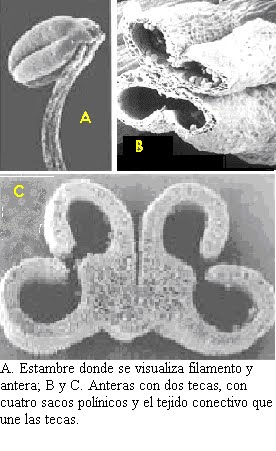
Estambres: El conjunto de los estambres de una flor constituye el androceo.
El estambre se compone de un filamento y de antera, y ésta, a su vez, consta de una porción estéril que está unida al filamento, el conectivo y de dos partes marginales, las tecas, en las que se contienen los sacos polínicos. Cada teca encierra dos sacos polínicos, por lo tanto, en las angioespermas un microesporofilo (estambre) contiene cuatro microsporangios. En el interior de estos sacos polínicos se encuentra un gran número de células madre de las microsporas, cada una de las cuales origina por división reductiva (meiosis) cuatro granos de polen.
Los granos de polen tienen forma esférica o elipsoide, y están protegido, salvo raras excepciones, por la exina y la intina. Sólo la intina, que esta formada de pectina y de celulosa, se desarrolla posteriormente constituyendo el tubo polínico en el interior del estilo. Para el paso de éste existen el exina puntos de salida preformados, en forma de pliegues de membrana más delgada, lunares redondeados o incluso poros abiertos o provistos de un opérculo.
Formación del gametofito masculino o microgametofito:
Ya en el saco polínico el núcleo del grano de polen (micróspora), al principio unicelular se divide en la proximidad de la pared de dicho grano y origina además de un núcleo vegetativo mayor, otro núcleo llamado generativo. En estas condiciones el grano de polen esta listo para pasar al estigma, donde la intina constituye el tubo polínico; en el ápice de éste se localiza el núcleo vegetativo y el cual además de dirigir la formación del tubo polínico posee una función trófica extrayendo nutrientes de las células del estilo. Dentro del tubo polínico el núcleo generativo se divide en dos núcleos espermáticos.
El estambre se compone de un filamento y de antera, y ésta, a su vez, consta de una porción estéril que está unida al filamento, el conectivo y de dos partes marginales, las tecas, en las que se contienen los sacos polínicos. Cada teca encierra dos sacos polínicos, por lo tanto, en las angioespermas un microesporofilo (estambre) contiene cuatro microsporangios. En el interior de estos sacos polínicos se encuentra un gran número de células madre de las microsporas, cada una de las cuales origina por división reductiva (meiosis) cuatro granos de polen.
Los granos de polen tienen forma esférica o elipsoide, y están protegido, salvo raras excepciones, por la exina y la intina. Sólo la intina, que esta formada de pectina y de celulosa, se desarrolla posteriormente constituyendo el tubo polínico en el interior del estilo. Para el paso de éste existen el exina puntos de salida preformados, en forma de pliegues de membrana más delgada, lunares redondeados o incluso poros abiertos o provistos de un opérculo.
Formación del gametofito masculino o microgametofito:
Ya en el saco polínico el núcleo del grano de polen (micróspora), al principio unicelular se divide en la proximidad de la pared de dicho grano y origina además de un núcleo vegetativo mayor, otro núcleo llamado generativo. En estas condiciones el grano de polen esta listo para pasar al estigma, donde la intina constituye el tubo polínico; en el ápice de éste se localiza el núcleo vegetativo y el cual además de dirigir la formación del tubo polínico posee una función trófica extrayendo nutrientes de las células del estilo. Dentro del tubo polínico el núcleo generativo se divide en dos núcleos espermáticos.


Hojas carpelares: Las hojas carpelares o simplemente carpelos forman junto con los primordios seminales el gineceo de la flor.
En las angioespermas, los carpelos se sueldan siempre en uno o varios recipientes cerrados, los ovarios. Cada ovario contiene uno o varios primordios seminales y hacia arriba remata con un estigma, que sirve para la captura y la germinación del polen, por lo que suele ser papiloso o mucilaginoso-pegajoso. Entre ovario y estigma con frecuencia se desarrolla un estilo que es la prolongación estéril del ovario; su parte interior sirve para conducción y nutrimento del tubo polínico.
Los primordios seminales, que brotan de la placenta y reciben alimento por medio de haces conductores, se hallan en general situados claramente sobre las hojas carpelares, que por esta razón, los carpelos, son considerados como macroesporofilos. Los primordios seminales poseen tegumentos o capas protectoras llamados primina y secundina, los mismos rodean una estructura más o menos ovoidea, masiva y parenquimática, llamada nucelo, excepto en un sitio de apertura llamado micrópila.
El nucelo es diploide y es el verdadero macrosporangio, se diferencia en primer lugar durante el curso de su desenvolvimiento la célula madre de las macrósporas.
El primordio seminal está unido a la placenta por un pie o funículo que es recorrido por un haz vascular hasta la zona de la calaza o chalaza, que es la base del nucelo.
Formación del gametofito femenino o macrogametofito:
En la célula madre de la macróspora se realiza la división reductiva (meiosis), y en consecuencia se forman cuatro células haploides, por lo regular dispuestas en fila. De ellas sin embargo, sólo una, generalmente la inferior es la célula funcional, por lo tanto, es la única macróspora superviviente. A partir de ésta macróspora se formará el macrogametofito, en un primer momento es uninucleada y luego experimenta tres divisiones consecutivas (mitosis), primero en dos núcleos, luego en cuatro y finalmente en ocho núcleos. Con ello queda preparada la formación del gametofito femenino. De los ocho núcleos, tres de ellos se diferencian en el extremo inferior en las células denominadas antípodas que deben desempeñar una función fisiológica de tipo trófico. De los tres núcleos del extremo superior cercano a la micrópila, dos de ellos constituyen las sinérgidas, células acompañantes del tercer núcleo que deviene en la ovocélula. Los dos núcleos restantes corresponden a los núcleos polares.
En las angioespermas, los carpelos se sueldan siempre en uno o varios recipientes cerrados, los ovarios. Cada ovario contiene uno o varios primordios seminales y hacia arriba remata con un estigma, que sirve para la captura y la germinación del polen, por lo que suele ser papiloso o mucilaginoso-pegajoso. Entre ovario y estigma con frecuencia se desarrolla un estilo que es la prolongación estéril del ovario; su parte interior sirve para conducción y nutrimento del tubo polínico.
Los primordios seminales, que brotan de la placenta y reciben alimento por medio de haces conductores, se hallan en general situados claramente sobre las hojas carpelares, que por esta razón, los carpelos, son considerados como macroesporofilos. Los primordios seminales poseen tegumentos o capas protectoras llamados primina y secundina, los mismos rodean una estructura más o menos ovoidea, masiva y parenquimática, llamada nucelo, excepto en un sitio de apertura llamado micrópila.
El nucelo es diploide y es el verdadero macrosporangio, se diferencia en primer lugar durante el curso de su desenvolvimiento la célula madre de las macrósporas.
El primordio seminal está unido a la placenta por un pie o funículo que es recorrido por un haz vascular hasta la zona de la calaza o chalaza, que es la base del nucelo.
Formación del gametofito femenino o macrogametofito:
En la célula madre de la macróspora se realiza la división reductiva (meiosis), y en consecuencia se forman cuatro células haploides, por lo regular dispuestas en fila. De ellas sin embargo, sólo una, generalmente la inferior es la célula funcional, por lo tanto, es la única macróspora superviviente. A partir de ésta macróspora se formará el macrogametofito, en un primer momento es uninucleada y luego experimenta tres divisiones consecutivas (mitosis), primero en dos núcleos, luego en cuatro y finalmente en ocho núcleos. Con ello queda preparada la formación del gametofito femenino. De los ocho núcleos, tres de ellos se diferencian en el extremo inferior en las células denominadas antípodas que deben desempeñar una función fisiológica de tipo trófico. De los tres núcleos del extremo superior cercano a la micrópila, dos de ellos constituyen las sinérgidas, células acompañantes del tercer núcleo que deviene en la ovocélula. Los dos núcleos restantes corresponden a los núcleos polares.

Fecundación
Cuando el tubo polínico a penetrado hasta el primordio seminal suelta su contenido, pero nunca directamente en la ovocélula, sino junto a ella, generalmente en una sinérgida, que entonces se desorganiza. Parece ser, que las sinérgidas provocan de alguna manera, tal vez segregando enzimas, la apertura del tubo polínico. El núcleo vegetativo de dicho tubo se desorganiza y se reabsorbe; en cambio uno de los dos núcleos espermáticos penetra en la ovocélula fecundándola, mientras el otro, adentrándose más se fusiona con los dos núcleos polares dando origen a la célula madre del endosperma (triploide).

De la ovocélula fecundada se origina el embrión, y del núcleo endospérmico, el endosperma. Ello ocurre de la siguiente manera: la ovocélula transformada en cigoto se rodea de una membrana de celulosa y luego se divide, a menudo por varios tabiques transversales, en una corta serie de células que cosntituyen el llamado proembrión. Solamente la célula o las células que ocupan uno de los extremos del proembrión y que se dividen por la aparición de tabiques longitudinales formarán más tarde el embrión propiamente dicho. Las células restantes constituirán el suspensor.
El suspensor empuja al embrión hacia el tejido nutricio que se esta formando, y muchas veces contribuye también a su nutrición. El suspensor se une a menudo al embrión mediante una célula llamada hipófisis, que también puede contribuir, aunque en pequeña escala a su desarrollo (formación de la pilorriza y a veces del ápice de la raíz).
Al principio es un cuerpo redondeado y pluricelular, divido primero en cuadrantes y en octantes después.
El suspensor empuja al embrión hacia el tejido nutricio que se esta formando, y muchas veces contribuye también a su nutrición. El suspensor se une a menudo al embrión mediante una célula llamada hipófisis, que también puede contribuir, aunque en pequeña escala a su desarrollo (formación de la pilorriza y a veces del ápice de la raíz).
Al principio es un cuerpo redondeado y pluricelular, divido primero en cuadrantes y en octantes después.

El embrión es en este momento un cuerpo relativamente masizo, mientras que el suspensor tiene la forma de un pedúnculo, de longitud variable, por lo general uniseriado y está unido a la pared del saco embrionario en el extremo micropilar.
Las células periféricas del embrión originarán las células del estrato epidérmico de los cotiledónes, del tallo hipocótilo y una parte de la pilorriza de la raíz principal. Las células profundas darán origen a todos los tejidos internos de la planta.
Los embriones de las mono y dicotiledóneas pueden ser de forma similar hasta la fase en que el cuerpo principal del embrión se hace globoso. Subsiguientemente, el embrión de las dicotiledóneas adquiere forma bilobulada, debido a la aparición de dos cotiledónes, mientras que el embrión de las monocotiledóneas se trasnforma en una estructura más o menos cilíndrica por extensión directa de un solo cotiledón.
La parte del ápice que queda en la hendidura situada entre los dos cotiledónes en desarrollo constituye la gémula o plúmula. En el extremo opuesto del eje hipocótilo se localiza la radícula, en cuyos tejidos intervienen las células del embrión y del suspensor.
Ya antes de que la ovocélula fecundada se divida por primera vez, suele dividirse el núcleo endospérmico, y comienza de esta manera a formar el endosperma, que en primer lugar sirve de nutrimento al embrión para luego extinguirse o para transformarse en un tejido reservante de la semilla, que sólo más tarde, al germinar aquélla, consumirá el embrión. El endosperma suele formarse de manera nuclear, es decir, su núcleo se divide en primer término en un gran número de núcleos libres. Sólo después de cierto tiempo se forman membranas celulares divisorias entre los núcleos libres, y finalmente, siguiendo un proceso que varía en sus detalles, toda la cavidad del saco embrional se rellena de células. En otros casos la división nuclear es seguida inmediatamente por la división celular. En las angiospermas, al contrario de lo que ocurre en las gimnospermas, la formación del endosperma es consecuencia de la fecundación. Es su formación secundaria es decir sólo se produce cuando lo requiere realmnete el desarrollo del embrión, que al principio se alimenta a partir de éste tejido nutricio normalmente triploide.
Las células periféricas del embrión originarán las células del estrato epidérmico de los cotiledónes, del tallo hipocótilo y una parte de la pilorriza de la raíz principal. Las células profundas darán origen a todos los tejidos internos de la planta.
Los embriones de las mono y dicotiledóneas pueden ser de forma similar hasta la fase en que el cuerpo principal del embrión se hace globoso. Subsiguientemente, el embrión de las dicotiledóneas adquiere forma bilobulada, debido a la aparición de dos cotiledónes, mientras que el embrión de las monocotiledóneas se trasnforma en una estructura más o menos cilíndrica por extensión directa de un solo cotiledón.
La parte del ápice que queda en la hendidura situada entre los dos cotiledónes en desarrollo constituye la gémula o plúmula. En el extremo opuesto del eje hipocótilo se localiza la radícula, en cuyos tejidos intervienen las células del embrión y del suspensor.
Ya antes de que la ovocélula fecundada se divida por primera vez, suele dividirse el núcleo endospérmico, y comienza de esta manera a formar el endosperma, que en primer lugar sirve de nutrimento al embrión para luego extinguirse o para transformarse en un tejido reservante de la semilla, que sólo más tarde, al germinar aquélla, consumirá el embrión. El endosperma suele formarse de manera nuclear, es decir, su núcleo se divide en primer término en un gran número de núcleos libres. Sólo después de cierto tiempo se forman membranas celulares divisorias entre los núcleos libres, y finalmente, siguiendo un proceso que varía en sus detalles, toda la cavidad del saco embrional se rellena de células. En otros casos la división nuclear es seguida inmediatamente por la división celular. En las angiospermas, al contrario de lo que ocurre en las gimnospermas, la formación del endosperma es consecuencia de la fecundación. Es su formación secundaria es decir sólo se produce cuando lo requiere realmnete el desarrollo del embrión, que al principio se alimenta a partir de éste tejido nutricio normalmente triploide.


14 de junio de 2010

Se hallan estas plantas, en lugares sombríos, diseminadas en todas partes del mundo con un número extraordinario de especies (cerca de 9000 especies), pero su principal desarrollo lo alcanzan en los trópicos, donde se manifiestan con una gran diversidad, desde formas enanas, reducidas, de pocos milímetros de tamaño hasta helechos arborescentes con tallos esbeltos de varios metros y frondes de hasta tres metros.
Estructura del esporofito del helecho

El esporofito muestra la división del cormo en tres órganos fundamentales: tallo o eje caulinar, hoja y raíz. A diferencia de los espermatófitos, los pteridófitos no poseen una raíz principal sino tan sólo raíces caulógenas que nacen de un rizoma subterráneo. Los helechos se caracterizan por sus grandes hojas más o menos pinnadas, que reciben el nombre de ¨frondes¨ y que están situadas a modo de roseta sobre un eje caulinar corto y por ello poco conspicuo. Por lo tanto, las hojas son los únicos órganos visibles fuera de la tierra (salvo en helechos arborescentes). Éstas son compuestas, y están formadas por un raquis y por los folíolos.

En las hojas se desarrollan también los esporangios. Sobre la base de la estructura de los esporangios podemos dividir a los Filicatae vivientes en dos subclases:
Las hojas que llevan esporangios pueden ser en todo parecidas a las hojas vegetativas (esporotrofofilos) o bien diferenciarse más o menos de los trofofilos asimiladores como esporofilos.
- Eusporangiados, cuyos esporangios están rodeados en la madurez por una pared pluriestratificada.
- Leptosporangiados, cuyos esporangios maduros poseen una pared monoestratificada; a esta subclase pertenece la mayoría de los helechos.
Las hojas que llevan esporangios pueden ser en todo parecidas a las hojas vegetativas (esporotrofofilos) o bien diferenciarse más o menos de los trofofilos asimiladores como esporofilos.

Los esporangios se disponen formando grupitos de configuración variada, llamados soros; se originan por protuberancia del tejido foliar, la placenta, y se hallan recubiertos en muchas especies, y así protegidos hasta la madurez, por una excrescencia membranosa del limbo de la hoja, el indusio. Cada esporangio deriva, por divisiones sucesivas, de una sola célula epidérmica, y constituye en la madurez una cápsula provista de un pedúnculo pluricelular, delgado; en esta cápsula, cuya pared está formada por un solo estrato de células, contiene un gran número de esporas.


En el interior de los esporangios se hallan las células madres de las esporas que por división reductiva producen cuatro esporas (meiósporas) cada una. Las esporas al igual que las de todos los pteridófitos (y espermatófitos) poseen un endosporio delicado que se dilata en el momento de la germinación y un exosporio más resistente que se desgarra cuando la espora germina.
La dispersión de las esporas se basa en un mecanismo de apertura especial: el anillo corre por el dorso y el ápice y avanza hasta la mitad de la cara ventral del esporangio; está constituído por una fila prominente de células con tabiques radiales e internos fuertemente engrosados; el anillo es un mecanismo de cohesión que (con ayuda de las células separadoras del estomio) provoca la dehiscencia del esporangio y el lanzamiento de las esporas.

Todos los helechos son isospóricos. De la espora germinante se origina el protalo, que a lo sumo alcanza algunos centímetros de longitud y vive solamente durante un tiempo limitado. El mismo presenta forma acorazonada y posee cloroplastos por lo cual es de color verde y puede realizar fotosíntesis. Se fija al sustrato por medio de rizoides.
Los anteridios y arquegonios se producen casi siempre en el mismo protalo y precisamente en la cara opuesta a la de incidencia de la luz, es decir, normalmente en la cara inferior, próxima al suelo húmedo.
Los anteridios se ubican próximos al vértice, entre los rizoides. Son órganos esféricos, abovedados, sin pedículo. El anteridio maduro está constituido por dos células anulares y una célula de cubierta (célula opercular), que encierran una célula central. De esta última se originan por división las células espermatógenas. Una vez desprendida la célula opercular quedan libres las espermátidas. Cada edspermátida produce un espermatozoide arrollado en tirabuzón y provisto de un penacho de flagelos; los espermatozoides son atraídos hacia los arquegonios por unos indicios de ácido málico.
 Los arquegonios se ubican cerca de la escotadura del protalo, son en número más reducidos que los anteridios. Tienen forma de jarro, con un vientre y un cuello. En el vientre se halla la ovocélula y la célula del canal del vientre. Entre las células de la pared del cuello se encuentra la célula del canal del cuello que es única y alargada. Las células del canal del cuello y del vientre estallan a consecuencia del hinchamiento de un mucilago que contienen y llenan el canal con una sustancia que se hincha mucho con el agua que entra, abriéndose así el cuello del arquegonio para que penetre el espermatozoide y fecunde a la ovocélula.
Los arquegonios se ubican cerca de la escotadura del protalo, son en número más reducidos que los anteridios. Tienen forma de jarro, con un vientre y un cuello. En el vientre se halla la ovocélula y la célula del canal del vientre. Entre las células de la pared del cuello se encuentra la célula del canal del cuello que es única y alargada. Las células del canal del cuello y del vientre estallan a consecuencia del hinchamiento de un mucilago que contienen y llenan el canal con una sustancia que se hincha mucho con el agua que entra, abriéndose así el cuello del arquegonio para que penetre el espermatozoide y fecunde a la ovocélula.Desarrollo del esporofito

La ovocélula fecundada se divide en cuatro cuadrantes por medio de dos paredes perpendiculares entre sí; una de estas células da lugar a la célula apical de la primera hoja; otra al ápice caulinar y una tercera al ápice radical. La cuarta célula desarrolla el pie, que permanece en el arquegonio y hace posible la alimentación inicial del embrión por parte del protalo.

4 de junio de 2010

Estructura del gametofito del musgo

El gametofito está formado por caulidios, filidios y rizoides, estructuras análogas a tallos, hojas y raíces respectivamente. Los rizoides fijan en el sustrato al caulidio foliado. En el extremo del caulidio y rodeado por pequeños filidios se desarrollan anteridios o arquegonios, generalmente varios de ellos mezclados con filamentos estériles llamados paráfisis. Los grupos de anteridios y arquegonios pueden aparecer distribuidos monoicamente, o sea ambos tipos en distintas ramas de la misma planta o bien en distribución dioica, o sea gametofitos poseedores de anteridios y gametofitos poseedores de arquegonios.
 ARQUEGONIO
ARQUEGONIOAl igual que en pteridófitos, tanto los esporangios como los gametangios son constantemente pluricelulares y se encuentran siempre envueltos por una cubierta de células estériles. Los macrogametangios llamados arquegonios, son órganos en forma de botella provistos de una pared constituída por una capa ordinariamente sencilla de células, en la cual se diferencian dos partes, llamadas vientre y cuello. La porción ventral encierra una célula central, grande, que se divide poco antes de la madurez y produce la ovocélula, y una célula del canal del vientre, situada en la base del cuello. A estas dos células siguen, en el cuello, las células del canal del cuello. Los briófitos poseen siempre un gran número de éstas; los pteridófitos sólo contienen una.
 ANTERIDIO
ANTERIDIOLos anteridios -microgametangios- son formaciones globosas o mazudas, cuya pared, formada por una sola capa de células encierra el tejido espermatógeno. Las pequeñas células espermatógenas, más o menos numerosas se dividen cada una en dos espermátidas que se desprenden de la masa de tejido en que se han originado y, sin otra división se transforman en sendos espermatozoides biflagelados.
La fecundación de los arquegonios ocurre en presencia de agua y por atracción quimiotáctica de los espermatozoides. El cigoto sufre primero una división transversal y forma en el extremo superior una célula apical bilateral cuyos segmentos orientados hacia los lados continúan dividiéndose y dan lugar a una cápsula esporífera, mientras que las células basales del embrión del esporofito desarrollan el denominado pie.
Aunque el esporofito presenta tejidos con cloroplastos, su actividad asimiladora no es suficiente. Por esta razón, depende para su nutrición del gametofito, en cuyos tejidos se fija por medio del pie que se desarrolla a modo de haustorios. En la parte distal del esporofito joven se producen rápidas divisiones celulares que dan lugar al desarrollo de la cápsula esporífera.
El esporofito joven esta rodeado al principio por las paredes del arquegonio, que durante un tiempo crecen junto con el esporofito. En la mayoría de los musgos acaban finalmente por desgarrarse en la parte basal. El fragmento mayor se eleva debido al intenso crecimiento de la seta, y forma la caliptra que recubre total o parcialmente la cápsula esporógena. En el interior de esta cápsula encontramos a las células madres de las esporas, células que aún son diploides. Tras pasar por dos estapas de división cada una de estas células madres de las esporas produce cuatro meiosporas. Las esporas poseen una pared doble: un endosporio delicado queda rodeado por un exosporio más resistente que se desprende en el momento de la germinación.
Aunque el esporofito presenta tejidos con cloroplastos, su actividad asimiladora no es suficiente. Por esta razón, depende para su nutrición del gametofito, en cuyos tejidos se fija por medio del pie que se desarrolla a modo de haustorios. En la parte distal del esporofito joven se producen rápidas divisiones celulares que dan lugar al desarrollo de la cápsula esporífera.
El esporofito joven esta rodeado al principio por las paredes del arquegonio, que durante un tiempo crecen junto con el esporofito. En la mayoría de los musgos acaban finalmente por desgarrarse en la parte basal. El fragmento mayor se eleva debido al intenso crecimiento de la seta, y forma la caliptra que recubre total o parcialmente la cápsula esporógena. En el interior de esta cápsula encontramos a las células madres de las esporas, células que aún son diploides. Tras pasar por dos estapas de división cada una de estas células madres de las esporas produce cuatro meiosporas. Las esporas poseen una pared doble: un endosporio delicado queda rodeado por un exosporio más resistente que se desprende en el momento de la germinación.

 En el ápice de la cápsula se suele desarrollar un complicado mecanismo de apertura. Las capas externas de tejido se convierten en un opérculo. Éste se apoya en un anillo de células que en el momento de la madurez de la cápsula mueren y despiden el opérculo al variar su estado de turgencia. Por debajo del opérculo se encuentra a menudo el denominado peristoma, formado por uno dentículos que pueden abrir o cerrar la cápsula gracias a sus movimientos higroscópicos.
En el ápice de la cápsula se suele desarrollar un complicado mecanismo de apertura. Las capas externas de tejido se convierten en un opérculo. Éste se apoya en un anillo de células que en el momento de la madurez de la cápsula mueren y despiden el opérculo al variar su estado de turgencia. Por debajo del opérculo se encuentra a menudo el denominado peristoma, formado por uno dentículos que pueden abrir o cerrar la cápsula gracias a sus movimientos higroscópicos. 

La espora, haploide, al germinar produce un filamento verde muy ramificado, positivamente fototrópico, el protonema. Sus primeras células son muy ricas en cloroplastos y presentan tabiques transversales perpendiculares; más tarde las células se vuelven probes en cloroplastos y los tabiques se hacen en parte oblicuos. Cuando la intensidad luminosa es suficiente se forman en el protonema las yemas productoras de gametofitos, como protuberancias primeramente laterales generalmente de la célula basal de una de las ramas laterales del protonema. Después de la separación de una o dos células pediculares, a partir de la célula terminal ensanchada se origina una célula apical trigonal cuya segmentación da lugar a la pequeña planta foliosa del musgo. Cuando se producen muchas yemas de este tipo los musgos se disponen a menudo formando céspedes.

Imagen de protonema al microscópio con yemas.

Imagen al microscópio de protonema.

31 de mayo de 2010

{kind=link}
{kind=link}
{kind=link}
Suscribirse a:
Comentarios (Atom)